
1. Introductions
Legumes are affected by biotic factors such as insects, molds, bacteria, and viruses. These plants can produce many different molecules in response to the attack of phytopathogens. Protease inhibitors (PIs) are proteins produced by legumes that inhibit the protease activity of phytopathogens. PIs are known to reduce nutrient availability, which diminishes pathogen growth and can lead to the death of the pathogen. PIs are classified according to the specificity of the mechanistic activity of the proteolytic enzymes, with serine and cysteine protease inhibitors being studied the most. Previous investigations have reported the efficacy of these highly stable proteins against diverse biotic factors and the concomitant protective effects in crops, representing a possible replacement of toxic agrochemicals that harm the environment.
Legumes (Fabaceae) are the second most important crops in the world, after the grass or Gramineae family (Poaceae) [1]. Legume seeds are widely used for direct food use because of their high nutritional content (Table 1) [2] and bioactive compounds (dietary fiber, polyphenols, flavonoids, phytosterols, resistant starch, and micronutrients like essential minerals and vitamins) [3,4,5].
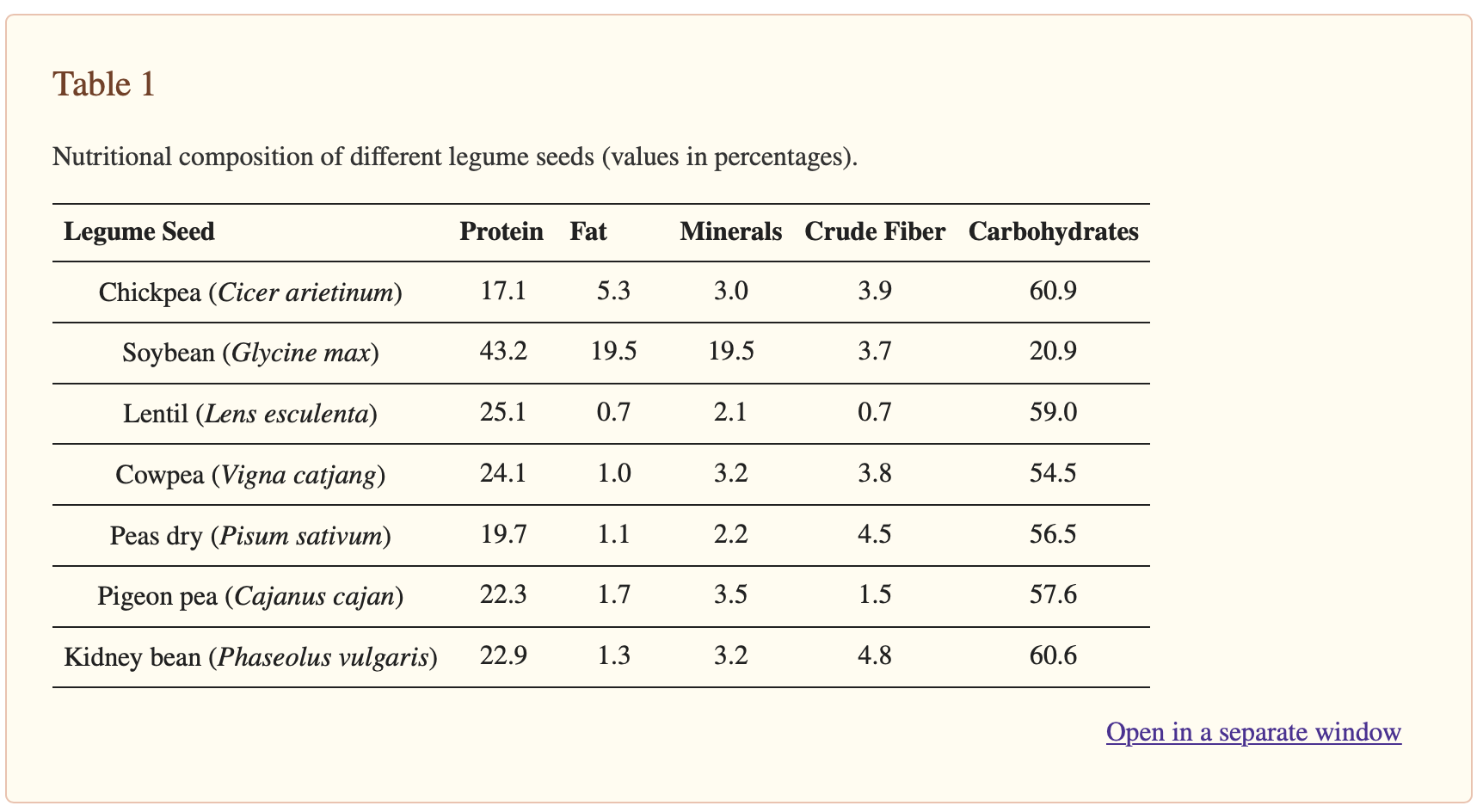
The Table was modified from Kamboj et al. [2].
Like many other relevant crops, legumes are exposed to different environmental stresses that affect their yield. These stress factors can be abiotic or biotic. The latter include fungi, bacteria, viruses, nematodes, and herbivorous insects [6,7,8]. Legumes are especially attacked by different species of the Fusarium genus (F. oxysporum, F. solani, F. udum, and F. virguliforme). These molds are responsible for the formation of vascular wilt and root rot and, therefore, lead to reduced crop yield. Likewise, many phytopathogenic bacteria, such as Pseudomonas and Xanthomonas, negatively affect legume plants by causing leaf blights and spotting. Viruses are transmitted by insects and may cause different symptoms on different host plants. Viruses primarily predispose legumes to other pathogen infections [9]. The nematodes that are known to have the most significant impact on legumes are root-knot (Meloidogyne spp.) and cyst nematodes (Heterodera spp. and Globodera spp.); they cause substantial damage in legume crops [10]. The insects, Callosobruchus chinesis, and C. maculatus, are the most important pests of legumes because they damage the seeds, causing a loss of dry matter weight, nutritional quality, and germination or viability [11,12]. These biotic factors severely affect legume crops, which can lead to significant economic losses and reduced world food production.

Currently, the use of agrochemicals is the principal way to eliminate, control, or prevent the attack of biotic agents. However, due to their toxicity and danger to human health, there is a necessity to replace them with non-toxic or less toxic products.
Legume plants synthesize and accumulate molecules in response to biotic stressors, known as antinutritional factors (ANFs). ANFs are compounds that reduce the bioavailability of nutrients through the inhibition of enzymes involved in digestion or by chelating minerals during pathogen infestations. Importantly, some ANFs are known to have toxic effects in living organisms when consumed at high doses [5]. Despite the presence of ANFs, the use of legumes as human food sources is not limited by the presence of these compounds. Several effective methods are utilized to inactivate or reduce the activity of ANFs [13]. ANFs are classified as protein- and non-protein-based compounds. Several studies have shown their potential benefits, including their use as biopesticides, anti-cancer agents, weight control, immune-modulators, and hypocholesterolemia regulators; additionally, there are other essential health benefits [14,15].
In response to a pathogen attack, legumes produce protein-based ANFs called pathogenesis-related (PR) proteins. According to Van Loon [16], PR proteins are those proteins that are not or only at basal concentrations detectable in healthy tissues, but for which accumulation at the protein level has been demonstrated upon pathological conditions and related situations in at least two or more plant–pathogen combinations. Van Loon [17] also introduced the term “inducible defense-related proteins” for PR proteins.
PR proteins are classified into PR-1 through PR-17 and act against pathogens by different mechanisms such as cell wall degradation (glucanase, chitinase), oxidative activity (peroxidase, oxalate oxidase), protease inhibitor, protein degradation (endoprotease), membrane permeabilization (thaumatin-like, defensin, thionin, lipid-transfer protein), degradation of RNA (ribonuclease-like), and other unknown mechanisms [18]. PR proteins that act as protease inhibitors (PIs) are classified as PR-6. These proteins inhibit the activity of protease enzymes in the pathogens; therefore, they are unable to feed on the amino acids present in the plant. In the particular case of legumes, they are capable of producing a great variety of PIs. The natural defense mechanisms of these plants can be exploited to avoid or decrease the use of toxic agrochemicals. For this reason, in the present review, we discuss the potential use of these proteins as biopesticides to control biotic stresses in crops of economic importance.
2. Legume Responses to Pathogen Attack
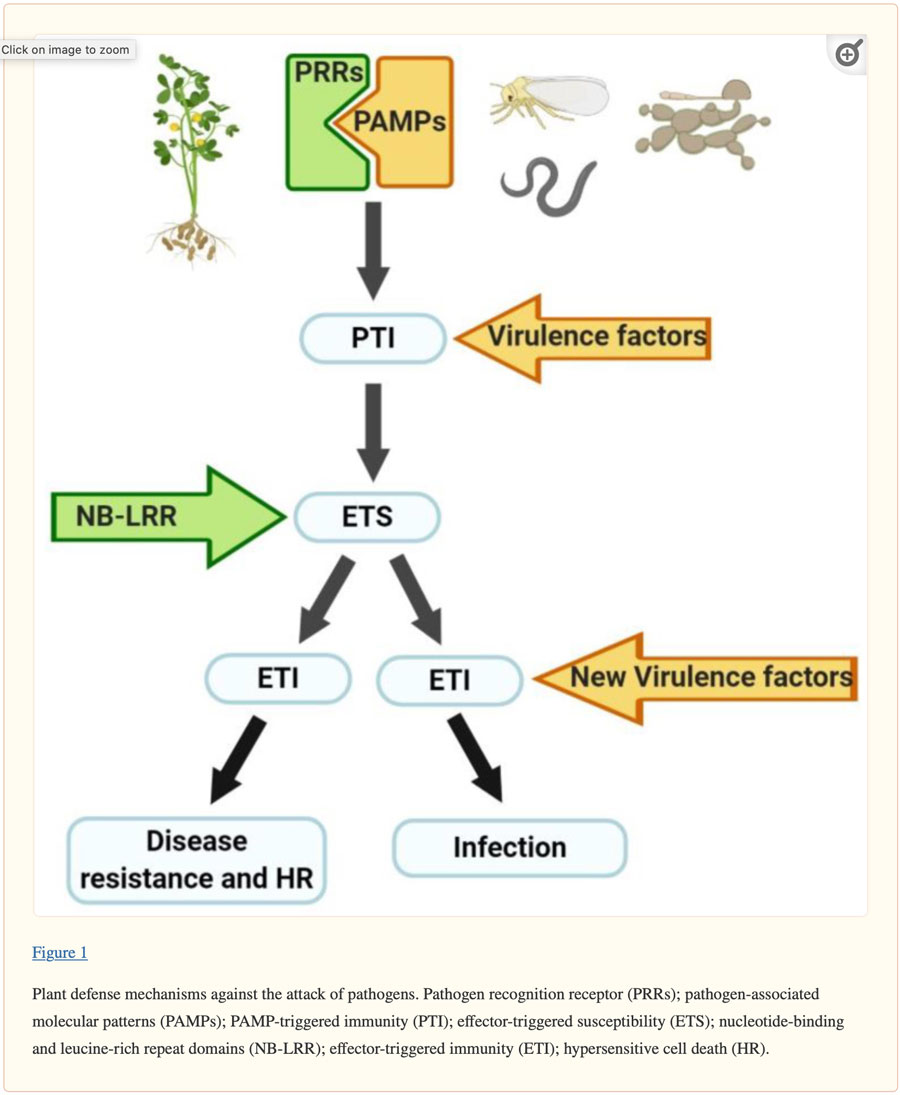
Plants have developed different defense mechanisms in response to biotic stressors. As shown in Figure 1, when a pathogen is present, the plant uses cellular proteins, called pathogen recognition receptor (PRRs), to recognize inherent molecules of the pathogen, called pathogen-associated molecular patterns (PAMPs). PAMP-triggered immunity (PTI) is the result of this recognition process and is used by plants to initiate a response to cease or ameliorate pathogen colonization. Some pathogens are capable of producing effector molecules (virulence factors) that interfere with PTI, resulting in effector-triggered susceptibility (ETS). Plants can synthesize some NB-LRR (nucleotide-binding and leucine-rich repeat domains) proteins that recognize the pathogen effector, which in turn develops the effector-triggered immunity (ETI). ETI results in disease resistance and typically hypersensitive cell death (HR) at the infection site. Successful pathogens can avoid ETI by diversifying recognized effectors, or acquiring new effectors that eliminate NB-LRR proteins; this can lead to colonization infection [19,20,21].
Specific hydrolytic enzymes, also called virulence factors or pathogen effectors, are used by pathogens to degrade the plant cell wall to get nutrients from the plant. The ability of the pathogen to invade a plant is determined mainly by enzyme activity. The activity of pathogen enzymes, proteases, has been directly correlated to the severity of plant disease, which indicates their important role in pathogen colonization. The proteases destroy the plant cell wall structural proteins, allowing the pathogen to obtain nutrients for its growth, as well as to evade plant defenses [22,23] (Figure 2a). The inhibition of pathogenic enzymes by the plant PIs constitutes a plant defense mechanism. The main mechanistic action of PIs is the loss of the ability to colonize the host plant and nutrient deficiencies, which in turn causes a decrease in the growth or the reproduction capacity and possibly the death of the pathogen (Figure 2b). The ability of plants to produce active PIs in response to a pathogen attack is an important feature related to plant survival [24].

3. Phytohormones and PIs in Legumes
Some PR proteins are directed by the salicylic acid (SA)-dependent signaling pathway, while others are controlled by ethylene (ET) or jasmonic acid (JA). The phytohormones SA, ET, and JA act as both signaling or inhibiting molecules [25]. On the other hand, abscisic acid (ABA), produced in the presence of a pathogen, is a crucial phytohormone in the octadecanoid pathway that leads the production of JA and the consequent expression of PIs in tomato plants [26]. Phytohormones play an essential role in plant immunity against pathogens, and they can act in different ways in diverse plant parts according to the invasiveness of the pathogen. Phytohormones interactions involve complex crosstalk that has been reviewed by several authors [27,28,29].
In the case of legumes, defense responses are also elicited by phytohormones. It has been observed that soybeans accumulate SA after insect damage, which induces the expression of PIs-specific genes. Likewise, the external application of JA in soybean plants also induces PIs genes [30]. Additionally, it has been demonstrated that phosphatidic acid (PA) increases rapidly in wounded and neighboring unwounded soybean leaves, which activates the production of proteins related to plant defenses [31]. Paudel and Bede [32] indicated an increase in the activity of PIs, in the leaves of Medicago truncatula, under attack by beet armywood caterpillars (Spodoptera exigua). Additionally, in this same model, it was found that the JA and ET pathways were also activated. On the other hand, SA and ABA were not induced by insects.
In a transcriptomic study conducted by Wang et al. [33], a gene encoding PIs was induced in the leaves of two lines of soybean treated with the common cutworm (Spodoptera litura fabricius). Genes related to the JA signaling pathways were up-regulated under these conditions. Gao et al. [34] identified the Acyrthosiphon kondoi resistance gene, AKR, in response to a bluegreen aphid A. kondoi attack on M. truncatula. The bluegreen aphid A. kondoi infestation did not induce the genes involved in octadecanoic or JA pathways in a susceptible line of the plant. However, the PIs genes were induced by the aphid infestation in a resistance line. During aphid attack of susceptible and resistant lines, genes involved in the SA pathway were induced, and it was suggested that activation of this pathway might be a general mechanism of aphid repellence, with limited effectiveness in susceptible hosts. The octadecanoic acid pathway plays an essential role in the induction of PIs. Importantly, this metabolite participates in the synthesis of JA, which induces expression of PIs. AKR, a resistance gene (R-gene) that encodes to an NB-LRR protein, mediates JA defenses [34,35]. Yamchi et al. [36] confirmed the role of the JA pathway and the expression of PIs in the resistance of M. truncatula against the phytopathogenic bacteria, Ralstonia solanacearum.
Kunkel and Brooks [37] concluded that both the JA and ET are typically associated with defenses against insects and necrotrophic pathogens and that SA and JA/ET defense pathways are commonly mutually antagonistic.
